 对照组 (P<0.05)。墨吉明对虾肝胰脏中超氧化物歧化酶(SOD)活力在48 h内呈下降趋势,且48 h SOD活力最小(148.90 U/mg prot),而48 h后SOD活力逐步上升。在胁迫时间24~96 h范围内,肝胰脏中溶菌酶活力随着亚硝酸盐胁迫时间的延长而逐渐升高,96 h达到最大值(0.62 U/ml)。墨吉明对虾血清蛋白含量、肝胰脏SOD活力和溶菌酶活力在3个取样时间点均存在显著差异(P<0.05),而对照组在3个取样时间点差异不显著(P>0.05),而在3个取样时间点对照组血清蛋白含量和2种酶的活力均未达到显著差异(P>0.05)。[结论]墨吉明对虾对亚硝酸盐的安全浓度达到11.48 mg/L,证实该虾类对亚硝酸盐具有较高的耐受性。当水体中亚硝酸盐浓度超过安全浓度时,会导致墨吉明对虾的死亡。
对照组 (P<0.05)。墨吉明对虾肝胰脏中超氧化物歧化酶(SOD)活力在48 h内呈下降趋势,且48 h SOD活力最小(148.90 U/mg prot),而48 h后SOD活力逐步上升。在胁迫时间24~96 h范围内,肝胰脏中溶菌酶活力随着亚硝酸盐胁迫时间的延长而逐渐升高,96 h达到最大值(0.62 U/ml)。墨吉明对虾血清蛋白含量、肝胰脏SOD活力和溶菌酶活力在3个取样时间点均存在显著差异(P<0.05),而对照组在3个取样时间点差异不显著(P>0.05),而在3个取样时间点对照组血清蛋白含量和2种酶的活力均未达到显著差异(P>0.05)。[结论]墨吉明对虾对亚硝酸盐的安全浓度达到11.48 mg/L,证实该虾类对亚硝酸盐具有较高的耐受性。当水体中亚硝酸盐浓度超过安全浓度时,会导致墨吉明对虾的死亡。
关键词墨吉明对虾;亚硝酸盐;半致死浓度;免疫因子
中图分类号S949文献标识码A文章编号0517-6611(2015)21-160-04
海洋甲壳动物免疫易受到多种环境因子(如温度、盐度、污染物等)的胁迫,其中亚硝酸盐作为水生态系统中氮代谢的中间产物,是水产养殖环境中主要的污染物之一。亚硝酸盐不仅能抑制对虾的生长和代谢,而且会对机体内抗氧化酶活力、溶菌酶活力以及细胞呼吸爆发力等体液免疫因子产生严重影响。已有研究表明,亚硝酸盐可以改变罗氏沼虾(Macrobrachium rosenbergii)肝胰脏中超氧化物歧化酶(SOD)活力,降低斑节对虾(Penaeus monodon)血蓝蛋白携氧能力,引起凡纳滨对虾(Litopenaeus vannamei )体内溶菌酶、血清蛋白含量变化等。此外,亚硝酸盐对其他水生动物的免疫因子也存在影响。例如,罗非鱼(Tilapia)体内溶菌酶活力随着亚硝酸盐浓度的升高而降低;三角帆蚌(Hyriopsis cumingii)的免疫因子随着亚硝酸盐浓度的升高均下降;中华绒螯蟹(Eriocheir sinensis)幼体随着亚硝酸盐浓度升高以及胁迫时间的延长,SOD活力呈现下降趋势。因此,控制养殖水体中亚硝酸盐浓度对增强对虾免疫水平和提高存活率至关重要。
墨吉明对虾(Fenneropenaeus merguiensis),俗称大虾、大明虾,隶属节肢动物门(Arthropoda)、甲壳纲(Crustacea)、对虾属(Penaeus),广泛分布于我国广东、广西沿海地区,此外在菲律宾等地也有分布。随着凡纳滨对虾养殖规模的扩大,病害发生、种质退化以及进口亲虾价格攀升等问题严重影响着对虾养殖产业和产量,因此众多水产工作者开始寻求可以替代凡纳滨对虾的品种。墨吉明对虾作为本地品种,其抗病性较强、生长速度较快、价格较进口凡纳滨对虾低,因此备受业内人士的关注。目前,国内外对墨吉明对虾的研究主要集中在性成熟与胚胎发育、年龄与增长、基因表达、资源调查等方面,而关于亚硝酸盐胁迫对墨吉明对虾毒性及免疫因子的影响的研究尚未见报道。因此,笔者采用静水生物毒性试验法研究了亚硝酸盐对墨吉明对虾的毒性作用,并且探究了高浓度亚硝酸盐胁迫下墨吉明对虾体内血清蛋白含量、超氧化物歧化酶(SOD)和溶菌酶活力的变化情况,旨在揭示墨吉明对虾适合生存的亚硝酸盐浓度,为墨吉明对虾的大规模健康养殖提供理论依据。
1材料与方法
1.1试验材料
试验于2014年7月在广东海洋大学无脊椎动物工程中心进行,试验所用墨吉明对虾由广东海洋大学水产学院自行繁育获得,挑选体色正常、体质健壮的墨吉明对虾进行试验,其体长为(51.54±10.24)mm,腹节长为(29.14±8.05)mm。试验容器采用经消毒的100 L塑料白桶。墨吉明对虾暂养7 d,每天换水1/2,暂养期间连续充气,每天投喂东腾颗粒3~4次,每次投喂量占对虾体质量的30%~40%。试验用水为经过消毒、曝晒后的海水,海水温度为26~28 ℃,盐度23‰。亚硝酸盐浓度采用亚硝酸钠(分析纯)配制,购于广东光华有限公司。
1.2亚硝酸盐急性毒性试验
亚硝酸盐浓度根据毒性预试验结果按等对数间距设置6个浓度梯度(33.20、52.79、8393、133.45、212.19、337.38 mg/L),同时设置对照组(亚硝酸盐浓度<0.02 mg/L),每组均设置3个平行,各放入20尾墨吉明对虾。试验期间,每天换水1/2,且保持各组浓度不变,供给充足氧气,不投喂饵料,观察并记录24、48、96 h对虾死亡情况,及时清除死亡个体。
1.3免疫指标的测定
另外选取健康的20尾墨吉明对虾,饲养在亚硝酸盐浓度为144.99 mg/L的白桶中,分别在24、48、96 h各取3尾对虾进行取样。用消毒过的1 ml注射器从墨吉明对虾心脏中抽取血液,并加入到预冷的抗凝剂中,血液与抗凝剂的体积比为1∶1,抗凝剂采用Gross等方法配制。将抽取血液8 000 r/min离心5 min后,取上清用于血清蛋白含量的测定。另外选取墨吉明对虾肝胰脏,将其放入灭菌后的研磨器中,加入1 ml生理盐水进行研磨,研磨后的匀浆液置于4 ℃条件下保存,用于SOD和溶菌酶活力的测定。
血清蛋白含量测定采用考马斯亮蓝法,在595 nm处测定样品吸光值大小,并对照标准蛋白的吸光值,计算出蛋白浓度。超氧化物歧化酶(SOD)活力测定采用南京建成试剂盒,每毫升反应液中SOD抑制率达50%时所对应的SOD量为1个SOD活力单位。溶菌酶活力的测定采用南京建成试剂盒,在规定条件下于450 nm处每分钟使吸光度值降低0001为1个酶活力单位。
1.4数据处理
采用直线内插法计算24、48、96 h亚硝酸盐急性毒性半致死浓度(LC50),并根据求出的96 h LC50按照以下公式计算安全浓度(SC):
SC=0.1×96 h LC50。
使用SPSS13.0统计软件对测定免疫指标进行单因素方差分析,试验结果均以平均值±标准差表示,P<0.05表示差异显著,P<0.01表示差异极显著。
2结果与分析
2.1亚硝酸盐急性毒性试验结果
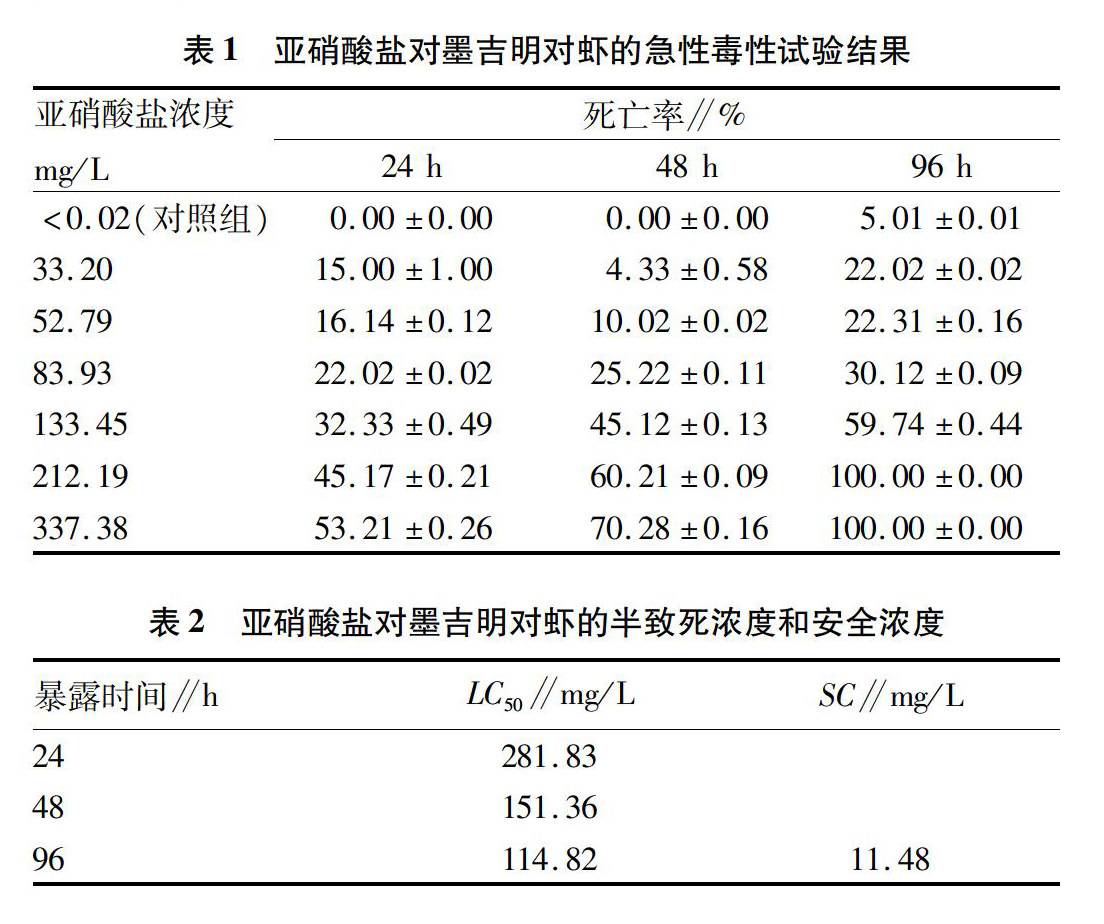
在亚硝酸盐急性毒性试验过程中,对照组中所有试验对虾食欲旺盛,很少出现空胃现象,且游动速度较快,受到外界刺激后弹跳力强;亚硝酸盐胁迫组墨吉明对虾几乎不摄食,空胃现象普遍,游动速度较慢,受到外界刺激后弹跳无力。此外,中毒的对虾尾部、足部会微微发红,死亡后体色呈现灰白色。由表1可知,对照组在24 h和48 h胁迫时间内并无个体死亡,96 h开始出现死亡。在同一胁迫时间,随着亚硝酸盐浓度的增加,对墨吉明对虾产生毒性效应增强,死亡率升高,且不同浓度组之间死亡率存在显著差异(P<0.05);同一亚硝酸盐浓度下,随着胁迫时间的延长,亚硝酸盐在墨吉明对虾体内富集,毒性效应增强,个体死亡数目逐渐增加,且不同胁迫时间下死亡率差异达显著水平(P<0.05)。由表2可知,墨吉明对虾24、48、96 h的半致死浓度分别为281.83、151.36和114.82 mg/L,其安全浓度为11.48 mg/L。
2.2亚硝酸盐胁迫对墨吉明对虾免疫因子的影响
从图1可以看出,对照组血清蛋白含量显著高于试验组血清蛋白含量(P<0.05)。随着胁迫时间的延长,血清蛋白含量呈先逐渐升高后逐渐下降的趋势,48 h血清蛋白含量最高为0.24 mg/ml。而对照组中24~96 h墨吉明对虾血清蛋白含量并未发生显著变化(P>0.05),基本维持在0.5~0.6 mg/ml范围内。
从图2可以看出,对照组中墨吉明对虾肝胰脏SOD活力显著高于试验组(P<0.05),且24~96 h对照组SOD活力的变化不显著(P>0.05),基本维持在350~400 U/mg prot范围SOD。24~48 h试验组SOD活力被抑制,SOD活力逐渐下降;在48~96 h SOD抑制解除,SOD活力逐渐上升,亚硝酸盐胁迫48 h后墨吉明对虾SOD活力最小,为148.90 U/mg prot。
从图3可以看出,高浓度亚硝酸盐在3个胁迫时间对墨吉明对虾肝胰脏的溶菌酶活力有影响。在试验胁迫时间范围内(24~96 h),溶菌酶活力呈逐渐升高趋势,96 h溶菌酶活力达最大值(0.62 U/ml),且亚酸盐胁迫下3个时间点之间溶菌酶活力差异显著(P<0.05)。对照组中溶菌酶活力变化不显著(P>0.05),活力维持在0.13~0.16 U/ml。亚硝酸盐胁迫48~96 h时实验组溶菌酶活力显著高于对照组溶菌酶活力(P<0.05)。
3讨论
3.1亚硝酸盐对墨吉明对虾的毒性作用
对虾养殖水环境中亚硝酸盐主要来源于微生物、虾类代谢废物、残余饵料以及死亡个体的分解作用,随着对虾养殖规模的不断扩大,水体中亚硝酸盐积累量增多,积累的亚硝酸盐主要以NO-2和HNO2的形式进入对虾体内,并对机体产生毒害作用。该试验结果表明,在相同浓度条件下,随着胁迫时间的延长,亚硝酸盐毒性作用增强,墨吉明对虾死亡率升高;在相同胁迫时间下,随着亚硝酸盐浓度的增大,墨吉明对虾的死亡率也呈现逐渐升高的变化趋势。对凡纳滨对虾、中国对虾(Penaeus chinensis)、日本沼虾(Macrobrachium nipponense)、克氏原鳌虾(Procambarus clarkia)等研究发现,随着亚硝酸盐浓度的升高和胁迫时间的延长,对虾死亡率逐渐增加。不同种类对虾的96 h半致死浓度和安全浓度见表3。该试验结果表明,在盐度为21‰条件下,体长约为51.54 mm墨吉明对虾的96 h半致死浓度和安全浓度分别为11482和11.48 mg/L。与同等规格的斑节对虾、凡纳滨对虾相比,墨吉明对虾的亚硝酸盐半致死浓度和安全浓度较高。墨吉明对虾对亚硝酸盐的耐受能力较高,其原因可能是由于物种差异导致体内的代谢机能不同,从而影响对亚硝酸盐的耐受能力。
3.2亚硝酸盐对墨吉明对虾血清蛋白含量影响
甲壳动物血清蛋白中含有大量的血蓝蛋白,因此血清蛋白具有载氧功能,此外血清蛋白还与免疫密切相关。该试验结果表明,随着亚硝酸盐胁迫时间的延长,墨吉明对虾血清蛋白含量先升高后下降,且显著低于正常水平(对照组)(P<0.05)。这与黄翔鹄、张亚娟研究结果相一致,在亚硝酸盐胁迫下凡纳滨对虾和日本沼对虾体内血清蛋白含量会被诱导升高,但显著低于对照组。在亚硝酸盐胁迫下,墨吉明对虾血清蛋白含量低于对照组的原因可能包括以下方面:①随着胁迫时间的延长,部分蛋白质被加速代谢为氨氮;②血淋巴中多种免疫酶因亚硝酸盐作用而受到抑制,导致表达量下降,造成血清蛋白含量低于正常水平。墨吉明对虾血清蛋白含量虽低于对照组,亚硝酸盐胁迫24~48 h后血清蛋白含量有所升高,其原因可能是亚硝酸盐引起墨吉明对虾呼吸障碍,机体通过提高血清蛋白含量来增加携氧能力,抵抗外界刺激,因此在短时间内墨吉明对虾体内蛋白含量会增加。但是,若亚硝酸盐胁迫时间过长,机体免疫系统被破坏,无法调节自身机能,血清蛋白含量则出现下降趋势,携氧能力减弱,对虾窒息死亡。
3.3亚硝酸盐对墨吉明对虾SOD活力的影响
超氧化物歧化酶是水生动物体内一种重要的抗氧化酶,在清除超氧化自由基O-2、防止机体氧化损伤、维持机体中氧自由基平衡等方面发挥重要作用。因此,水生动物体内SOD活力能够较好反映机体的免疫能力。该试验结果表明,亚硝酸盐胁迫下墨吉明对虾SOD活力显著低于对照组,且差异达显著水平(P<0.05)。对罗氏沼虾和中华绒螯蟹的研究也发现,在亚硝酸盐胁迫下SOD活力显著降低(P<0.05),并低于对照组。该试验结果表明,试验组墨吉明对虾SOD活力变化呈现一定规律性,即先下降后升高,这与黄翔鹄研究亚硝酸盐胁迫下凡纳滨对虾SOD活力变化有所不同。产生的原因可能是亚硝酸盐胁迫条件下,墨吉明对虾呼吸爆发产生大量的活力氧自由基,短时间内抑制SOD活力,但随着胁迫时间的延长,机体通过自身调节,SOD表达量增多,SOD活力增强,氧自由基被清除,机体产生较弱的抗氧化能力,因此48 h后SOD活力上升。墨吉明对虾在亚硝酸盐胁迫下,短时间内通过提高自身的SOD活力,增强机体的抗氧化能力;然而,凡纳滨对虾SOD活力显著下降,并无短暂升高,导致亚硝酸盐的耐受力低于墨吉明对虾。
3.4亚硝酸盐对墨吉明对虾溶菌酶活力的影响
溶菌酶是广泛存在于水生动物血清、吞噬细胞中的一种碱性蛋白,能够对革兰氏阳性菌起到破坏并消除的作用。因此,它在一定程度上能够反映环境因素改变对水生动物免疫水平的影响。该试验结果表明,在亚硝酸盐胁迫下墨吉明对虾溶菌酶活力逐渐上升,且48~96 h溶菌酶活力高于对照组。黄翔鹄和丁美丽等在对凡纳滨对虾、中国对虾的研究中发现,随着亚硝酸盐胁迫时间的延长,溶菌酶活力显著降低,与该试验结果不同。墨吉明对虾具有一定的耐受性,会调节自身相关的免疫机制,因此在亚硝酸盐胁迫下逐渐出现上升趋势,且趋势变化显著(P<0.05)。已有研究表明,亚硝酸盐胁迫能够降低凡纳滨对虾和罗氏沼虾的免疫力,增加副溶血弧菌的易感性,造成对虾死亡率增大。墨吉明对虾溶菌酶活力会在亚硝酸盐胁迫下升高,致使对虾的抗病能力增强,降低致病菌的易感性,提高存活率。因此,墨吉明对虾与其他品种对虾相比具有更强的耐亚硝酸盐能力。
综上所述,亚硝酸盐胁迫下墨吉明对虾免疫因子的变化,反映了对虾在不同时间段对亚硝酸盐的响应,而其诱导机制则有待进一步研究。该试验条件下,墨吉明对虾各免疫因子水平会随胁迫时间的延长而提高,增强对虾抗亚硝酸盐能力,因此与其他品种对虾相比亚硝酸盐的安全浓度较高。
参考文献
[1] 艾春香,李少菁,王桂忠,等.几种环境因子对虾蟹类非特异性免疫的影响[J].海洋通报,2007,26(5):102-107.
[2] JENSEN J B.Nitrite disrupts multiple physiological functions in aquatic animals[J].Comparative Biochemistry and Physiology,2003,135(1):9-24.
[3] YIN X L,LI Z J,YANG K,et al.Effect of nitrite on growth and molting of Penaeus monodon juveniles[J].Fish Shellfish Immunology,2014,40(1):190-196.
[4] LE MOULLAC G,HAFFNER P.Environmental factors affecting immune responses in Crustacea[J].Aquaculture,2000,191(1):121-131.
[5] 王玥,胡义波,姜乃澄.氨态氮、亚硝态氮对罗氏沼虾免疫相关酶类的影响[J].浙江大学学报,2005,32(6):698-705.
[6] CHENG S Y,CHEN J C.Hemocyanin oxygen affinity,and the fractionation of oxyhemocyanin and deoxyhemocyanin for Penaeus monodon exposed to elevated nitrite[J].Aquatic Toxicology,1999,45(1):35-46.
[7] XIAN J A,WANG A L,CHEN X D,et al.Cytotoxicity of nitrite on haemocytes of the tiger shrimp,Penaeus monodon,using flow cytometric analysis[J].Aquaculture,2011,317(4):240-244.
[8] 黄翔鹄,李长玲,郑莲,等.亚硝酸盐氮对凡纳滨对虾毒性和抗病相关因子影响[J].水生生物学报,2006,30(4):466-471.
[9] 臧学磊.三种环境因子胁迫罗非鱼集体免疫力的变化及其对海豚链球菌易感性的影响[D].南京:南京农业大学,2010:9-10.
[10] 沈文英,谢优刚,邵秦.铵态氮、亚硝酸盐氮对三角帆蚌免疫酶活力的影响[J].浙江农业学报,2007,19(5):364-367.
[11] 洪美玲.水中亚硝酸盐和氨氮对中华绒螯蟹幼体的毒性效应及维生素E的营养调节[D].上海:华东师范大学,2007:55-71.
[12] 麦贤杰,黄伟健,叶富良,等.对虾健康养殖学[M].北京:海洋出版社,2009.
[13] PATIL P K,GOPAL C,PANIGRAHI A,et al.Oral administration of formalin killed Vibrio anguillarum cells improves growth and protection against challenge with Vibrio harveyi in banana shrimp[J].Letters in Applied Microbiology,2014,58(3):213-218.
[14] QURESHI N A,AMANAT Z.Reproductive biology and size at sexual maturity of penaeus merguiensis from the sonmiani bay lagoon,balochistan Pakistan[J].Journal of Animal and Plant Sciences,2014,24(2):503-511.
[15] MEMON A J,IKHWANUDDIN M,TALPUR A D,et al.Assessment of sperm viability,determination fertilization capacity and hatching rate by artificial Insemination of Banana Shrimp,Penaeus merguiensis(De Man,1888)[J].Research Journal of Applied Sciences,2011,6(3):174-178.
[16] MANE S,DESHMUKH V D.Age and growth studies of banana prawn,Penaeus merguiensis de Man from Maharashtra waters[J].Journal of the Marine Biological Association of India,2011,53(2):184-188.
[17] 王芸,李健,李吉涛,等.pH胁迫对中国明对虾抗氧化系统酶活力及基因表达的影响[J].中国水产科学,2011(3):556-564.
[18] 钟振如,李辉权,闽信爱,等.南海北部硇洲岛周围水域中的墨吉对虾资源[J].水产学报,1992,14(4):277-285.
[19] GROSS P S,BARTLETT T C,BROWDY C L,et al.Immune gene discovery by expressed sequence tag analysis of hemocytes and hepatopancreas in the Pacific White Shrimp,Litopenaeus vannamei,and the Atlantic White Shrimp,L.setiferus[J].Developmental & Comparative Immunology,2001,25(7):565-577.
[20] 寇红岩,冼健安,郭慧,等.亚硝酸盐对虾类毒性影响的研究进展[J].海洋科学,2014,38(2):107-115.
[21] 王娟,曲克明,刘海英,等.不同溶氧条件下亚硝酸盐和氨氮对中国对虾的急性毒性效应[J].海洋水产研究,2007,28(6):1-6.
[22] WANG W N,WANG A L,ZHANG Y J,et al.Effects of nitrite on lethal and immune response of Macrobrachium nipponense[J].Aquaculture,2004,232(1):679-686.
[23] 於叶兵,陆伟,黄金田,等.亚硝酸盐和硫化物对克氏原螯虾幼虾的毒性效应研究[J].水生态学杂志,2011,32(1):111-114.
[24] 胡贤德,孙成波,王平,等.不同盐度条件下亚硝酸氮对斑节对虾的毒性影响[J].沈阳农业大学学报,2009,40(1):103-106.
[25] LEE S Y,LEE B L,SDERHLL K.Processing of an antibacterial peptide from hemocyanin of the freshwater crayfish Pacifastacus leniusculus[J].Journal of Biological Chemistry,2003,278(10):7927-7933.
[26] 张亚娟,王超,刘存歧,等.氨态氮和亚硝态氮对日本沼对虾酚氧化酶活力及血蓝蛋白含量的影响[J].水产科学,2010,29(1):31-34.
[27] BAIRD S,KELLY S M,PRICE N C,et al.Hemocyanin conformational changes associated with SDSinduced phenol oxidase activation[J].Biochimica et Biophysica Acta(BBA)Proteins and Proteomics,2007,1774(11):1380-1394.
[28] 洪美玲,陈立侨,孙新谨,等.亚硝酸盐急性胁迫对中华绒螯蟹幼体相关免疫指标和应激蛋白表达的影响[J].应用与环境生物学报,2011,17(5):688-693.
[29] 高明辉.VC、VE对亚硝酸盐胁迫下异育银鲫血液指标及抗氧化能力的影响[D].武汉:华中农业大学,2008.
[30] 陈家长,臧学磊,孟顺龙,等.亚硝酸盐氮胁迫对罗非鱼血清非特异性免疫酶活力的影响[J].生态环境学报,2012,21(5):897-901.
[31] 丁美丽,林林,李光友,等.有机污染对中国对虾体内外环境影响的研究[J].海洋与湖沼,1997,28(1):7-12.
[32] 葛红星,李健,陈萍,等.亚硝酸盐氮胁迫下凡纳滨对虾对副溶血弧菌的易感性[J].中国水产科学,2014(3):629-636.
[33] CHENG W,CHEN J C.Effects of environmental factors on the immune responses of freshwater prawn Macrobrachium rosenbergii and other decapodcrustaceans [J].J Fish Soc Taiwan,2002,29:1-19.







